

Knowledge
Glial Cells the forgotten half of the brain:
and their role in learning and memory
February 10, 2021
Successive learning involves the same neural circuits, which help to create memory.
Study of the anatomy of the nervous system and connective tissues. By Gustaf Retzius (1842–1919). Source: hagstromerlibrary.ki.se
By Pierre J. Magistretti
10/02/ 2021
·
- Share
What are neurons?
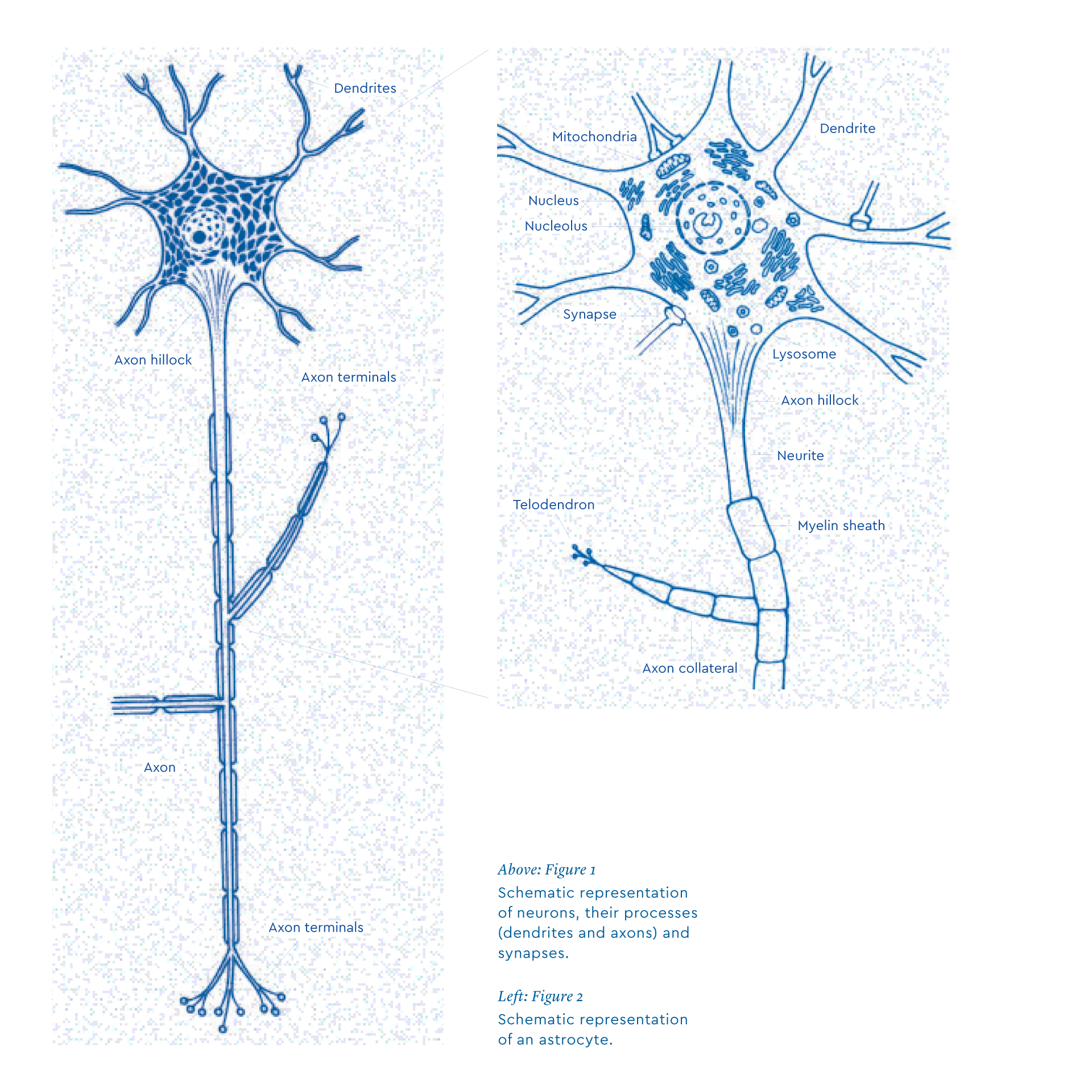
Neurons share many features with other cells in the human body, but they also have specific characteristics that underlie their functions. For example, they have several hundred extensions called dendrites. Dendrites receive inputs from other neurons at specialized points called spines. One of the extensions, which emerges from the cell body, is the axon. The axon ends in nerve endings, which come into contact with neighboring neurons. The human brain has around 85 billion neurons. As each neuron is connected to tens of thousands of other neurons, and sends about a thousand signals per second, it is estimated that the brain produces about a billion signals per second. Quite an amazing figure!
The space between each nerve terminal and the dendrites of the next neuron is narrow, about 200 angstroms, which is less than one millionth of a millimeter. This space is called a synapse. Neurons secrete from axon terminals a chemical substance called the neurotransmitter which is released in the synapse and binds to “receptors” located on the dendrites of the next neuron (Figure 1). It is therefore neurons that ensure the transfer of signals within the brain. There is, however, a wide variety of neurons, probably several thousands, inspiring the Spanish neuroscientist Santiago Ramón y Cajal to rather poetically refer to them as “the butterflies of the soul”.
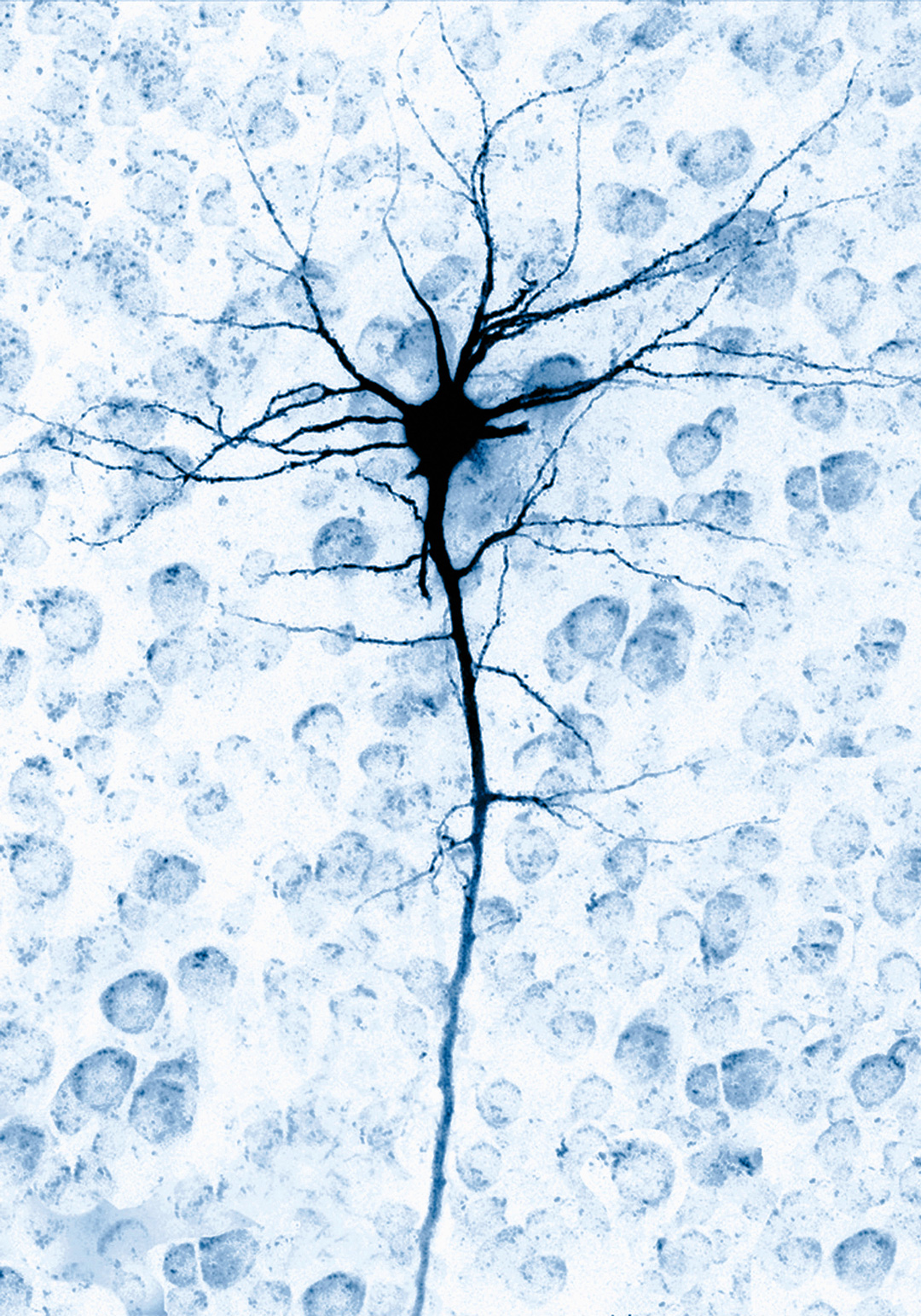
A single brain cell neuron.
What is the role of neurons in learning?
Neurons carry their messages in the form of electrical signals called action potentials. These are transmitted via the axon. Each action potential arriving at the end of a neuron (in the region of the synapse) triggers an activation or an inhibition response on the next neuron (on the opposite side of the synapse). The strength of these signals can vary.
Neurons are constantly changing. They are malleable enough to change the distribution of nerve connections as needed. During learning, sets of neurons change their configuration by multiplying synapses and making them more efficient to facilitate nerve transmission. This phenomenon is known as synaptic plasticity.
To ensure the transfer of new information, these new connections are made on demand. When information is repeated, the resulting neuronal signals are not randomly distributed. Instead, they tend to follow existing pathways. Everything happens as if the information has to go through the newly remodeled synapses repeatedly. As such, it is said that synapses are “strengthened”. Successive learning involves the same neural circuits, which help to create memory.
More on the architecture of the learning brain: Glial cells
At the frontiers of neuroscience research, it was recently discovered that neurons are not the only cells in the brain that are essential for learning. In certain species such as humans, there are at least as many non-neuronal cells, called glia.
Until recently, glial cells were considered to be a sort of “glue” that holds neurons together, a connective tissue of the brain. Over the past two decades, research on glial cells, particularly on a specific type called the astrocyte, has revealed the key role they play in brain function. Astrocytes provide energy for neurons and support synaptic plasticity during higher brain processes such as learning, memory and cognition.
The human brain contains slightly more glial cells than neurons, about 85 billion. The number of glial cells is slightly higher, more than 100 billion. The number and complexity of glial cells increase more with evolution and biological complexity than is the case with neurons. For instance, in a leech there is a single astrocyte for every 30 neurons. In an earthworm, there are six times more neurons than glia. In a rat the ratio of glia cells to neurons is 0.6:1, while in humans there are 1.5 glial cells per neuron. This suggests that glia density may be related to higher brain functions.
Glial cells were first described by the German neuropathologist Rudolf Virchow in 1856. He described a non-specific tissue, considered connective, that was interposed between the nerve cells and blood vessels. He concluded that this substance, which he called “neuro-glia”, was a kind of cement (“Nervenkitt”) that held the cells together. Towards the end of the nineteenth century, Camillo Golgi and colleagues proposed that glial cells play a role in the distribution of nutrients to neurons from the bloodstream.
Astrocytes take their name from the fact that they have many features that make them look like stars (in Greek, “astros” means “star”) (Figure 2). Astrocytes have specialized processes called end-feet that are in contact with blood capillaries, which in turn deliver energy to the brain (sugar and oxygen). A single astrocyte can contact hundreds of thousands of synapses. This means that astrocytes have the capacity to detect synaptic activity and couple it to the energy supply in the brain. Furthermore, astrocytes express receptors and reuptake sites that recognize neurotransmitters released at the synapse, particularly glutamate, which is the most common signal of neuronal activity released by 80% of synapses. When glutamate molecules are released at the synapse, they transmit information to other neurons and are quickly recorded by the astrocyte. When astrocytes detect synaptic activity, a series of metabolic processes is triggered that allows them to import sugar from the bloodstream and deliver energy to neurons. It turns out that astrocytes somehow “pre-digest” glucose (the main sugar in the blood), transforming it to lactate that can readily be used by neurons.
The classical view of neural communication mediated exclusively via synapses has expanded to include communication between neurons and astrocytes. In addition to lactate, astrocytes release other molecules that act on neurons and can even modulate synaptic activity. A new term has been coined to reflect this type of communication: gliotransmission (as opposed to neurotransmission). Astrocytes act as modulators of synaptic transmission. In some cases, glutamate released by a neuron acts not only conventionally on another neuron, but also on the receptors present in the astrocytic processes that surround the synapse. The astrocyte in turn releases glutamate which amplifies synaptic transmission between neurons. The astrocyte thus acts as a sort of “turbo charger” for synapses. Glutamate released by astrocytes can also act on an ensemble of neurons and thus synchronize their activity.
Emerging evidence on the role of glial cells in learning and memory
The conventional view of that, solely based on communication between neurons, is largely outdated. The neural and astrocytic networks engage in an intense dialogue via chemical signals, neurotransmitters, and gliotransmitters. This dialogue also highlights the role of astrocytes as integrators of neuronal activity.
Granted, neurons, through their remarkably efficient signaling capacities at the synapses, are essential to the transmission of information. The mechanisms of synaptic plasticity, which strengthen the transmission of information between groups of neurons, can be strengthened, and form a biological substrate for learning and memory. However, the understanding of memory exclusively based on neurons has been challenged, as more evidence indicates that glial cells, in particular astrocytes, also contribute to memory.
Astrocytes have a variety of properties that explain why they are critical players in memory and cognitive processes. For instance, they can sense and power synaptic activity. They produce molecules that can strengthen synaptic communication and they can synchronize the activities of groups of neurons involved in the learning process. Note that a single astrocyte can contact nearly 2 million synapses in human brains but only 100,000 in those of rodents. Interestingly, transplanting human astrocytes into the brain of newborn mice allows grafted animals to learn faster and to have increased memory capacity.
In a series of experiments carried out in my laboratory and in collaboration with other laboratories over the last decade, we have demonstrated that signals released by astrocytes are necessary for memory consolidation. Thus when, with appropriate pharmacological tools, we hampered the transfer of lactate from astrocytes to neurons in the hippocampus —a brain region critical for memory— we were able to block memory consolidation of a learning task in rodents. This lactate originates from glycogen, the energy storage form of glucose contained in astrocytes. It is noteworthy that they are the only cells in the brain that have this property.
Initially we thought that the role of lactate in memory consolidation was that it was acting as an energy substrate for energy-demanding tasks that underlie synaptic plasticity. However, we discovered that the lactate released by glial cells involved another property of lactate, namely the increase in the expression of genes necessary for synaptic plasticity.
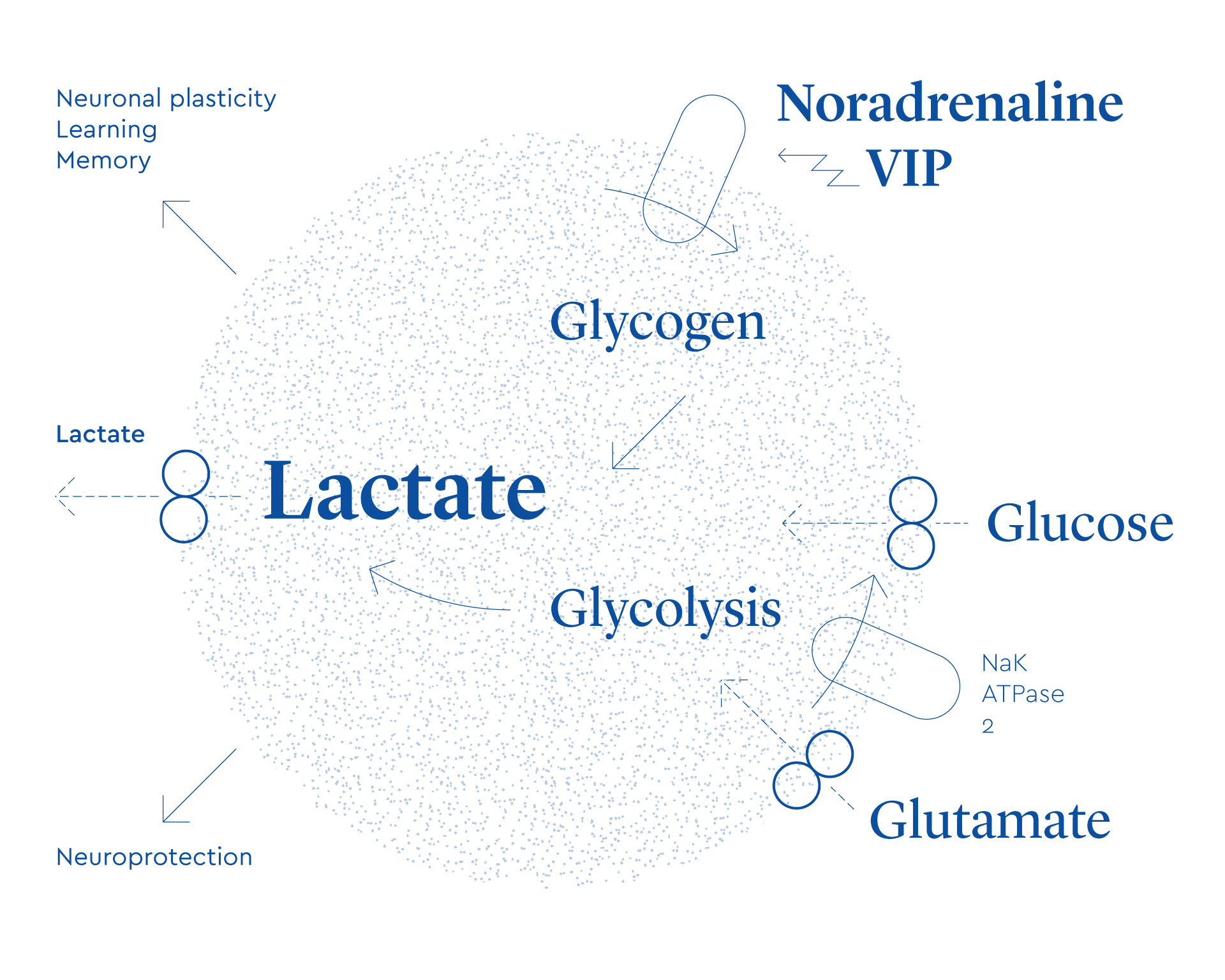
Schematic representation of neuron-glia interactions, highlighting the role of astrocytes-derived lactate in physiological functions.
Where are we and what is next?
These results demonstrated for the first time that a glia-derived molecule is necessary for memory consolidation. This is a much more auspicious role than “brain glue”. Similarly, we revealed that lactate, a molecule long considered as a metabolic end-product, plays a vital role in memory function (Figure 3).
We are now working to discover drugs that could promote the formation of lactate from astrocytes to increase memory performance in normal, as well as in pathological conditions such as dementia. Perhaps with time, such memory performance can be applicable to learning, especially the learning of complex tasks. Even for lower-level tasks, enhanced memory performance could improve the automaticity of task performance in order to release short-term memory for higher-level tasks.
There is no doubt that much research still remains to be done to identify all the cellular and mechanisms of memory consolidation, but those efforts must not only consider neurons but also glial cells as potential sites of action.